昆虫の移動の研究で注目されるハーモニックレーダー
トンボ(昆虫)の移動の観察においても各種の技術が用いられていて、その技術は日々進化していると言えます。Maggioraら (2019) は近年、昆虫の生態・行動学研究の場で用いられているハーモニックレーダー (Harmonic radar) を使ってイタリアでスズメバチの行動追跡を報じています。
これは一種の個体識別応答のトランスポンダーを背負った昆虫をミリ波レーダーで追跡するというものです。その技術自体は第二次世界大戦時に航空機の敵味方識別装置として開発されたものですが、現在これを小さなハチに載せてレーダーで追えるというのに驚かされます。ただこの技術はレーダーの出力が小さい(移動式で野外に簡易に設置するため)のでおのずと探知範囲が狭く、長距離移動するものは追いきれません。レーダー一基で周囲数百メートルといったところでしょうか。


スズメバチの移動経路▲ がレーダー、ピンクと黄色が巣 (Maggiora ら2019より)
先に、ヨーロッパマダラヤンマに発信器を付けて羽化後の行動を・・・といった研究例を紹介しましたが、今回の技術は少し異なっていて、トランスポンダーを使用しています。トランスポンダー自体は非電力型でレーダー電波を受けた時だけに自然発信するものなので、むしろレーダー側の性能向上が革新的であったともいえるかも知れません。先のヨーロッパマダラヤンマの例では、同時にヨツボシトンボの仲間にも装着していて、こちらは羽化後、5日ほど羽化場所からほど遠くない地域にそれらは滞留したことを確認していますから、成果が全くないわけではなかったのです。現在の技術を使えば、このヨツボシトンボについてはさらに興味深い事実がわかっていたものと思われます。
一方、アメリカの Wikelsk et al., (2006) は超小型発信器( 300mg、バッテリー内蔵式)を アメリカギンヤンマ Anax junius に接着して、そのトランスミッターからの電波を地上と受信アンテナ装着のセスナ2機によって12日間(バッテリ―の寿命がそのくらい)追跡しています。同時期にイタリアでおこなった実験は地上だけでの追跡でしたからやはりスケールが違う。いかにもアメリカらしい実験です。

その結果、移動はおおよそ3日に1回起きていて、方向は南、距離は3つのケースがあって、1つはやたらめったらに飛び、その1日の飛行距離は1-4km。もう1つは一定方向に8-12km。さらに一定方向に25-150kmと長距離を移動するものがあるそうです。押しなべて平均すると日当たり58kmであったとしています。これらの移動は鳥の渡りに非常に似ていて、彼らは関連性に注目しているようです(例えば昆虫にも日長と磁場を感じ取り相互の変化で移動するなど)。
日当たり58kmなら移動期間を単純に成熟期間と置き換えて、それに充てるならば、14日×58kmで730kmは移動可能です。案外日本産トンボにも、我々が考える以上にダイナミックな移動を行っている種類があるのかも知れません。
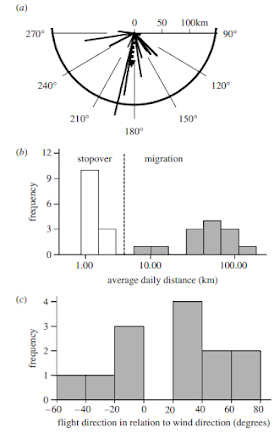
一番上が飛んだ方向、中が飛んだ距離と個体数(3つのグループに分かれる)
下が風の向きに対して飛行方向の関係 Wikelsk et al., (2006) より
トンボ科や生殖行動に入ったヤンマやヤマトンボなどの行動にこうした追跡技術が応用されれば、これまでなかなか分からなかった雌雄の行動が見えて来るかも知れません。個人的にはマルタンヤンマのオスの行動がどうなっているのか、なんてぜひ研究してほしいと思います。
幼虫の生育
さて、ヨーロッパマダラヤンマが西ヨーロッパ~北アフリカに広く分布していて、南ヨーロッパや北アフリカの発生状況は西ヨーロッパとはかなり異なることを前項で紹介しました。今回はこれまで公表された文献をもとに幼虫の生育についてみてみたいと思います。

Schiel et al., (2016)より
上に掲げた表はヨーロッパ産ルリボシヤンマ属(皇帝ギンヤンマも入ってます)の幼虫生育のデータの一部です。これを見ると、ヨーロッパマダラヤンマは他種(複数年、羽化までかかる種)に比べて孵化時期がかなり早く、また羽化までの齢数が少ないのが分かります。
これに対して日本のマダラヤンマの場合は、1967年に曽根原さんが小諸市で詳細に調べていて孵化が4月23日、9齢を経て7月23日に羽化したと報告しています。また、4月30日までに複数が孵化して8月17日までに羽化し、それらのヤゴ期間は3-3.5ヶ月(92-107日)であったとしています。上の表からヨーロッパマダラヤンマとの違いは、1ヶ月程度マダラヤンマの孵化は遅れるが、生育期間はほぼ同じといえそうです。しかし、なぜ緯度が高いドイツなどの西ヨーロッパで孵化時期が日本のマダラヤンマより早まるのでしょう?水温も低いと思われるのに幼虫生育速度も速く、7月には羽化が始まるのはどういうことなのでしょう。そう思って、北海道各地と長野県上田市の生息地さらにドイツ西部の生息地の気温を比較してみました。そうすると、意外にも上田市とドイツ西部における幼虫期間の気温差はほとんど無いことが分かりました(幼虫期間はグレーの帯で示してあります)。そうすると、単純に孵化時期の違いがそのまま羽化時期の差になっているに過ぎないのでしょうか(北海道は後ほど)。


一方、Schaller(1972)は越冬卵の温度環境を変えた時の孵化状況を観察しています。それがこの上のグラフです。非常に分かりずらいグラフなんですが、ヨーロッパマダラヤンマは20℃定温の状態 ⓸ だと産卵後75日で孵化が始まります。これに対して10℃定温状態のものを産卵後35(①)、70(②) および105日 (③) に20℃に加温した場合、孵化の同期率が70日後に加温したものが最も高かったとしています。
さらに幼虫の齢数についても、ヨーロッパマダラヤンマは産卵された卵は同じ環境の下で、すでに幼虫がたどる齢数が決まっているのだと言います。

この図も分かりずらいのですが、aはあご幅と齢数の関係で、b は生育期間と齢数の関係。縦軸は齢数、a の横軸はたぶんミリで、b は日です。確かにこの図からは生育環境条件が同じなら、孵化した時点で齢数は決まっているのだといえるかもしれません。生育度から齢数が決まるのではないようですね。
彼らは、本来ヨーロッパマダラヤンマは地中海性気候のもとで繁栄した種で、次第に北方に分布を広げた(温暖化の影響もあるか?)、と考えているようで、一定期間の低温状態に卵が置かれ、その後高温(発育可能な温度)になることで、胚発生が一斉に起きて、さらに短い発育期間で羽化する。そのタイミングは実験で低温期間が70日必要だったというのです。

一方、Munoz-Pozoら(1996)はスペイン南部で時期を変えて幼虫採取をおこなったところ、冬季の1~2月に最も小さな頭幅の幼虫が採集され、4月のはじめから5月にかけて終齢幼虫が採取されたと報じて、スペイン南部では一部が幼虫越冬であることを確認しました。こうなると水温の変化が知りたくなります。

池沼の水温の周年変化を深さ(ヤゴの生息環境に合わせた)ごとに計測した論文は意外に少なく、各県の水産試験場がおこなっている(場合もある)調査では水深が深すぎたりして、岸部のヨシ原内部などのお目当ての浅い水深でのデータがなかなか見つかりません。水性昆虫の分野では当然そうしたデータはあるのでしょうが、、。
Sternberg (1994) はスイス国内のトンボ生息地の沼地の日内水深別温度変化をグラフで示しています。これを見ると結構トンボが生息しそうな水深では温度の逆温度成層帯ができていることが分かります。夜間かなり温度が下がっても、水温はほぼ一定で、変化も緩やかなことが分かります。多分、月変化でもそうなのでしょう。
これらをひっくるめた上で、今度は南スペインのと西部ドイツのヨーロッパマダラヤンマの生活環を比較してみます。

上に示した図でみると、かなり両者は異なることが分かります。スペインの夏は暑く、平均気温がどうも18-19℃以上の暑さになると、成虫の行動が抑制されるようだという事の他に、地中海性気候は夏期の降水量が少なく場合によっては生息地が干上がる可能性が出てきます。だからこの時期に生殖行動をおこなって産卵してしまうことを夏眠(適当な言葉が思いつかなかったので)して避けているのだという研究者がいます。アキアカネのようにこの時期の個体は性成熟しないのでしょうか?今回、それはわかりませんでした。これに対してドイツでは成虫期間に平均気温が18-19℃に達せず、夏眠は起きないのかも知れません。ドイツでは晴れの日なら気温が10℃でオスが飛翔して、13℃でメスが産卵に飛来するとするデータがあるそうです。ちょっと日本では考えられないような低温でも活動するのですね。
一方、本種の発育零点が何℃かは分かりませんが、南スペインでは思ったほど水中温度は気温に比べて低温にはならないらしいことと、10月に産下された卵は1月までに約90日ありますから、その間15℃あたりの水温であれば十分この間に胚発生するのかもしれません。
つづく

.gif)
.gif)












.jpg)




























